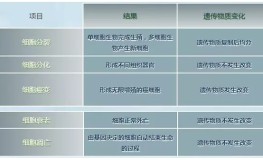
滩羊以其营养丰富、无膻味、肉质细嫩而著称。 肉的色泽稳定性通常由肌红蛋白(Mb)、血红蛋白和一些微量有色代谢物维持。在充分放血的肌肉中,Mb含量占总色素含量的75%左右,是影响宰后肉色泽稳定性的主要因素。 氧化磷酸化(OXPHOS)系统由线粒体内膜电子传递链(ETC)和ATP合成酶(复合物V)组成,是物质在线粒体内氧化释放的能量通过呼吸链供给ADP与无机磷酸合成ATP的偶联反应。 蛋白质组学技术是能够揭示细胞/组织生长、发育、代谢等生命现象本质变化的分子生物学筛选技术。 利用蛋白组学技术可以从分子水平上阐明肉品质在贮藏过程中的变化机制。
宁夏大学食品科学与工程学院的王金霞、魏志宝、罗瑞明*等人 采用iTRAQ蛋白质组学技术研究贮藏0、4、8 d滩羊肉中差异蛋白的表达变化,筛选与氧化磷酸化相关差异蛋白,鉴定0~4 d和4~8 d两个贮藏阶段共有的差异丰度蛋白质,采用Cytoscape软件鉴定核心网络;将所筛选的差异蛋白标注于氧化磷酸化通路上,从通路水平揭示氧化磷酸化调控滩羊肉色泽稳定性的机制,旨在为滩羊肉宰后肉品色泽稳定性控制提供一定理论支持。
P<0.05),贮藏4~8 d其活性显著上升(P<0.05),表明抗氧化酶体系清除自由基的能力开始减弱。 贮藏期间SOD活性随着贮藏时间的延长呈显著下降趋势(P<0.05),表明SOD清除超氧阴离子自由基的能力变弱。 GSH-Px活性在贮藏期间显著下降(P<0.05),表明GSH-Px将过氧化氢转化为水和分子氧的能力变弱。

P<0.05),这是因为内源抗氧化酶的活力随贮藏时间的延长而降低,导致ROS逐渐积累增加。

如图3所示,贮藏过程可以很好地区别样本,相同贮藏期的样本之间区别不显著,而不同贮藏期的样本之间区别显著,以上结果说明本实验样品收集合理,并且蛋白质组学数据可靠,可以满足后续分析的需要。
如图4所示,4 d对比0 d组记为4/0 d组,8 d对比4 d组记为8/4 d组,共鉴定出164 个丰度差异显著蛋白质。其中4/0 d组共鉴定出71 个丰度差异显著蛋白质,53 个显著上调,18 个显著下调;8/4 d组共鉴定出93 个丰度差异显著蛋白质,其中11 个显著上调,82 个显著下调。


利用GO注释对所筛选出的差异丰度蛋白质从生物过程(BP)、分子功能(MF)、细胞组分(CC)3 个角度对其功能进行生物信息统计。由图5a可知,4/0 d组的差异丰度蛋白质富集到406 个BP、98 个CC、95 个MF;由图5b可知,8/4 d组的差异丰度蛋白质富集到699 个BP、85 个CC、168 个MF。


由图6a可知,滩羊宰后贮藏0~4 d期间,差异丰度蛋白主要参与的BP包括肌肉收缩、肌肉系统过程、系统过程的调节、小分子代谢过程以及ATP代谢过程等;CC主要包括肌节、细胞外泌体、胞外细胞器、细胞外膜结合细胞器、横纹肌细丝等;MF主要包括维生素结合、谷胱甘肽转移酶活性、电子载流子活性等过程。滩羊屠宰后贮藏初期肌肉收缩、前体代谢和能量的产生、ATP代谢过程等发生变化,这些变化可能引起线粒体损伤,导致细胞内能量无法正常交换,能量代谢活动发生紊乱,致使MetMb无法被进一步还原,导致肌肉中3 种Mb的含量与存在形式发生改变,最终对滩羊肉色泽稳定性产生影响。
由图6b可以看出,滩羊宰后贮藏4~8 d期间,差异蛋白主要参与的BP包括ATP代谢过程、核苷二磷酸化、嘌呤核苷单磷酸代谢过程等;CC主要包括细胞外膜细胞器、细胞外泌体、膜结合囊泡、肌纤维、收缩纤维等;MF主要包括抗氧化物活性、细胞骨架蛋白结合、MHCII类蛋白复合物结合等过程。滩羊屠宰后贮藏后期,差异表达蛋白质涉及过氧化物酶活性、横纹肌收缩和ATP代谢过程等反应过程,这些代谢过程的变化可能影响内源氧化还原酶系统的活性,促使肌细胞线粒体能量代谢受到影响,引起肌肉组织收缩,直接影响3 种Mb的存在状态和含量,最终对滩羊肉色泽稳定性产生负面影响。
将3 个贮藏期所筛选的164 个差异丰度蛋白质进行KEGG通路注释统计,如图7所示,4/0 d组共显著富集于16 条KEGG通路,8/4 d组共显著富集于18 条KEGG通路。

滩羊宰后贮藏0~4 d期间,差异蛋白主要参与碳代谢、代谢途径、2-氧代羧酸代谢、半胱氨酸和蛋氨酸代谢以及心肌收缩等通路;宰后贮藏4~8 d期间,差异蛋白主要参与氨基酸合成、糖酵解/糖异生、磷酸戊糖途径、氧化磷酸化、HIF-1信号通路等。宰后初期,所筛选的差异蛋白并未明显富集于氧化磷酸化通路的根本原因可能是同一个差异蛋白可富集于不同的信号通路,例如氧化磷酸化复合物亚基COX5A、COX4I1及COX5B等都显著富集在心肌收缩通路中。以上代谢过程可能通过影响宰后肌细胞线粒体能量代谢活动从而影响高铁肌红蛋白还原酶(MetMbase)体系,最终影响滩羊肉色泽稳定性。同时,所筛选的差异蛋白可以显著富集到氧化磷酸化代谢通路,且随贮藏时间的延长其上的相关酶会发生改变,MetMbase系统又受氧化磷酸化的调控。因此,本研究将以氧化磷酸化通路中关键蛋白的差异变化为重点,初步阐明氧化磷酸化通路上关键蛋白对滩羊肉色泽稳定性产生的影响。
通过Venn网站()筛选3 个贮藏期共有的差异蛋白,如图8所示,4/0 d组与8/4 d组共有的差异蛋白为28 个,并将这28 个差异蛋白列于表1中。通过String网站()对28 个共有的差异蛋白进行PPI网络分析,如图9所示,该网络中PPI的富集
P值为1.0×10 -16,聚类系数为0.556。共有34 个节点数,78 条边位于该互作网络中,表明24 个差异蛋白之间具有明显的互作关系。
将PPI网络导入Cytoscape 3.9.1软件中并采用MCODE筛选该PPI网络中与氧化磷酸化相关的核心网络,结果如图10所示。核心网络上共有8 个的差异蛋白在氧化磷酸化通路中,分别为细胞色素c氧化酶亚基5A(COX5A)、COX2、ATP合酶亚基α(ATP5A1)、ATP合酶亚基O(ATP5O)、ATP合酶亚基D(ATP5H)、琥珀酸脱氢酶复合铁硫亚基B(SDHB)、ADP/ATP转位酶(SLC25A4)和细胞色素c(CYCS),其中ATP5A1、ATP5O、ATP5H参与线粒体中氢离子的转运;COX5A指导COX的正确装配,维持COX结构的稳定,COX2为细胞色素c氧化酶亚基2;SDHB负责将电子从琥珀酸盐转移到泛醌;SLC25A4负责催化ADP和ATP在膜上的交换。因此,认为这8 个上调表达的差异蛋白是影响滩羊宰后贮藏期间氧化磷酸化的核心差异蛋白。
为了将所筛选出的核心差异蛋白在氧化磷酸化通路中的变化进行可视化展示,以KEGG代谢途径作为媒介,将差异蛋白标注于氧化磷酸化通路中。由图11a可知,宰后贮藏0~4 d差异蛋白ATP5A1、ATP5O、ATP5H、COX5A、SDHB、SLC25A4和CYCS均上调表达,线也在贮藏初期上调表达,而COX2、COX3下调表达;Ndufv1、Ndufa8为泛醌氧化还原酶核心亚基,COX2、COX3、COX4、COX5A及COX5B均为细胞色素c氧化酶的核心亚基。由图11b可知,贮藏4~8 d氧化磷酸化通路上仍有多个差异蛋白上调表达,其中Ndufa13为NADH还原酶亚基,QCR8是泛醇-细胞色素c还原酶的亚基,负责编码低分子质量的泛醌结合蛋白。但宰后初期上调表达的COX4未发现明显变化,Ndufab1、COX5A显著下调表达。

如表2所示,贮藏0~4 d Mb表达量下调,这可能是因为机体供养方式的改变,促使Mb的初级结构发生改变,引起其开始下调表达,贮藏4~8 d期间,Mb为下调表达,这可能是因为随着贮藏时间的延长含量氧的降低使肌细胞内离子稳态失衡,造成线粒体功能受损,最终导致Mb表达量下降。滩羊背最长肌的Mb表达量在整个贮藏期呈显著下降趋势,这是因为Mb二级结构会随贮藏时间的延长发生分子重排,引起部分分子间的二硫键被破坏,导致Mb构象由规整向松散分布,致使Mb结构稳定性下降,最终导致滩羊背最长肌中的Mb表达量在整个贮藏期显著下降。

P<0.001),与ATP5O、ATP5H显著负相关(P<0.05),而其与COX2、COX5A呈高度显著正相关(P<0.001);ATP5A1与ATP5O、ATP5H、SLC25A4均极显著正相关(P<0.01),与SDHB、CYCS高度显著正相关(P<0.001),与COX2、COX5A呈极显著负相关(P<0.01);ATP5O、ATP5H与COX2、COX5A并无显著相关性,这可能是因为COX2、COX5A直接影响ATP5A1的表达量,从而间接对ATP5O、ATP5H产生影响;SDHB、SLC25A4、CYCS均与各指标显著相关。相关性分析结果表明,氧化磷酸化通路上的差异蛋白ATP5A1、ATP5H、COX2、COX5A、SDHB、SLC25A4和CYCS直接影响滩羊肉Mb的表达量,同时这些核心差异蛋白质也可能通过间接方式影响滩羊背最长肌的色泽稳定性。
ROS是具有活性化学性质的信号分子,能降低线粒体抗氧化能力,造成线粒体损伤,主要来自线粒体细胞。当机体受损或处于缺血缺氧状态时会产生过量的ROS,线粒体产生ROS的速率受线粒体内膜跨膜电位的调节。本研究中滩羊宰后贮藏期间滩羊背最长肌中的ROS含量显著上升,这是因为随着贮藏时间的延长内源抗氧化酶逐渐失活,不能及时清除机体内过量的ROS,导致ROS含量逐渐积累增加,从而导致氧化应激,最终引起细胞凋亡。
机体内的内源性抗氧化酶系统能够清除氧化应激产生的ROS。SOD是抗氧化酶系统中最重要的酶,主要负责催化超氧阴离子歧化成过氧化氢,CAT进一步将过氧化氢催化分解为水和氧气。马森提出GSH-Px通过激活谷胱甘肽介导了过氧化物、羟基等多种ROS的产生,从而维持了线粒体的稳定。本研究发现SOD、GSHPx活性随着贮藏时间的延长均显著下降,这说明内源抗氧化酶系统清除氧自由基的活力开始变弱。而CAT活性在贮藏0~4 d显著下降,但在4 d后却显著上升,这可能是因为同期SOD活力的上升催化产生了过多的过氧化氢,导致机体产生应激反应,致使CAT活力开始升高。
宰后滩羊肉的贮藏时间与肌细胞线粒体内能量代谢活动具有显著相关性,本研究发现不同贮藏阶段蛋白差异水平有显著变化,在贮藏期间显著上调或下调,且多个显著变化的差异蛋白可以富集于氧化磷酸化通路。滩羊宰后贮藏0~4 d,显著差异蛋白ATP5A1、ATP5O、ATP5H、COX2、COX5A、SDHB、SLC25A4及CYCS均富集于氧化磷酸化通路;贮藏4~8 d,除所筛选出的8 个核心差异蛋白外,差异蛋白Ndufab1、COX5A也富集于氧化磷酸化过程。虽然不同贮藏期鉴定的差异蛋白表达量并不相同,但这些差异蛋白在KEGG中均注释于能量代谢,这说明动物屠宰后,细胞内环境改变会引起组织内能量代谢相关酶亚基表达发生变化。线粒体蛋白质组在转录、翻译和活性等多个层面上存在协同作用,因此富集于氧化磷酸化通路上差异蛋白的上调/下调表达则说明肌细胞线粒体通过调节氧化磷酸化复合物的表达量和活性,保持正常的能量代谢活动,从而应对由于细胞内环境的改变而造成的线粒体损伤。蛋白质水平的上调表达很可能是由于线粒体受损而引起的离子稳态失衡、结构保护机制失效、抗凋亡功能失效等生理生化变化所致,蛋白质水平发生下调表达则预示着机体能量的持续耗竭,最终引起组织/细胞的程序性死亡。
所鉴定的差异蛋白在不同贮藏阶段呈现了上调或下调的表达趋势,细胞色素c氧化酶位于线粒体ETC的末端,其转录水平变化对酶活性具有调节作用,COX1、COX2及COX3具有酶催化活性,COX5A指导COX的正确装配,其中COX5A在贮藏初期上调表达,而在贮藏后期下调表达,这说明随着贮藏时间的延长肌细胞内环境的改变促使COX5A无法正确指导COX的装配,促使细胞色素c氧化酶活性发生改变,进而对氧化磷酸化进程产生影响,最终导致滩羊肉色泽稳定性受到影响。F型H+转运ATP酶广泛分布于线粒体内膜,是组织能量代谢的关键酶,负责利用呼吸链产生的电化学势能,改变蛋白质结构合成ATP。ATP5A1、ATP5O、ATP5H在贮藏初期均上调表达,在贮藏后期均下调表达,ATP酶亚基的蛋白质上调表达意味着宰后肌肉组织内发生应激反应,导致线粒体复合物V高效组装并引起其活性发生改变。ATP酶亚基的蛋白质水平下调表达意味随着贮藏时间的延长,细胞凋亡程序的启动使复合物V已无法正确装配,促使其活性开始下降。SDHB为复合物II亚基B,其具有结合金属离子和电子传递的功能对于阻止复合体II的电子泄漏起着非常重要的作用,可能介导呼吸链生物功能和调控细胞生长,SDHB在贮藏初期上调表达,并在贮藏后期下调表达,这可能是因为肌细胞内环境的改变促使复合物II亚基B的表达受到贮藏时间的调控,其表达量的下调表达会引起复合物II发生突变,促使其氧化效能增加,最终加速细胞死亡。
相关性分析结果表明,所筛选的8 个核心蛋白ATP5A1、ATP5H、COX2、COX5A、SDHB、SLC25A4和CYCS直接影响滩羊肉Mb的表达,并且这8 个核心蛋白均显著富集于氧化磷酸化通路中,因此推测所筛选出的8 个核心蛋白对滩羊肉色泽稳定性起重要调控作用,并且此过程是通过氧化磷酸化途径实现的。Mb衍生态在组织内存在的状态和相对含量为滩羊宰后肉色变化的内在机制,组织内的还原酶系统对Mb状态至关重要。而氧化磷酸化反应往往伴随着电子的转移,复合物I负责催化NADH脱氢生成NAD+,复合物II主要负责将琥珀酸催化生成延胡索酸,并将产生的电子最终传递给泛醌,复合物III主要负责将电子从辅酶Q的传递给细胞色素c,复合物IV主要负责将电子从细胞色素c转移到O2,将O2转化为水。已有研究表明,动物屠宰后机体细胞缺氧缺血,细胞内氧自由基的产生会引起肌细胞内离子稳态的失衡导致线粒体损伤,引起线粒体功能下降,促使氧化磷酸化复合物活性发生改变,导致MetMb还原酶系统受到影响,最终导致滩羊肉色稳定性变差。COX是联系低氧反应和代谢模式转化的重要纽带,它可以通过调节氧化磷酸化生成ATP,从而导致能量代谢方式发生转化,进而对滩羊被最长肌中的Mb表达量产生影响,最终影响滩羊肉色泽稳定性。
通过分析研究结果,发现宰后贮藏期间滩羊肉中内源抗氧化物酶系统的失效,导致ROS含量随贮藏时间的延长不断积累,引起肌细胞发生程序性凋亡,促使氧化磷酸化通路上ATP5A1、ATP5O、ATP5H、COX2、COX5A、SDHB、SLC25A4和CYCS等复合物亚基表达紊乱,引起线粒体复合物结构和功能发生改变,使氧化磷酸化进程受到阻碍,并进一步对肌肉中Mb存在形式造成影响,最终影响滩羊肉色泽稳定性。
采用iTRAQ技术分析鉴定出8 个差异蛋白质为影响滩羊肉色泽稳定性的核心蛋白质,分别为ATP5A1、ATP5O、ATP5H、COX2、COX5A、SDHB、SLC25A4和CYCS,这些蛋白质主要通过能量代谢过程和氧化还原过程影响滩羊肉色泽稳定性。宰后贮藏期间滩羊肉中抗氧化体系的失效导致ROS含量不断积累,破坏肌细胞线粒体完整性,促使氧化磷酸化通路上ATP5A1、ATP5O、ATP5H、COX2、COX5A、SDHB、SLC25A4和CYCS等复合物亚基表达紊乱,引起线粒体复合物结构和功能发生改变,使氧化磷酸化进程受到阻碍,对肌肉中MetMbase系统造成负向影响,最终影响滩羊肉色泽稳定性。
本文 《宰后贮藏期间氧化磷酸化调控滩羊肉色泽稳定性的机制》来源于《食品科学》2024年45卷第9期153-162页,作者:王金霞,魏志宝,陈雪妍,李荣,张倩,胡丽筠,罗瑞明。DOI:10.7506/spkx0425-250。点击下方 阅读原文 即可查看文章相关信息。
实习编辑:王雨婷 ;责任编辑:张睿梅。点击下方阅读原文即可查看全文。图片来源于文章原文及摄图网。
特别声明:以上内容(如有图片或视频亦包括在内)为自媒体平台“网易号”用户上传并发布,本平台仅提供信息存储服务。
女魔头Anna Wintour下岗|她是如何用职场PUA统治时尚圈37年的?
宣布成立“美国党”后,马斯克再发帖:应何时何地举行首次代表大会?会非常有趣
为什么买华为手机的人越来越少了? 这3个原因很线岁“星爷”,成中国第一人!穿鞋1米76,拿下世界顶级扣篮赛冠军
中国“不需要空调”的8座城市:夏季均温不超过25℃,晚上需要盖被子,风景绝美,人均500就能玩好
陕西3名初中生偷奔驰出车祸,定损43万保险拒赔,律师:监护人需承担主要赔偿责任
还在更新!撼讯推出RX 7600竞技V2版本:升级DP2.1、更大散热器